








方案详情
文
牡蛎降糖肽的结构表征及其协同花色苷的活性增效作用
方案详情

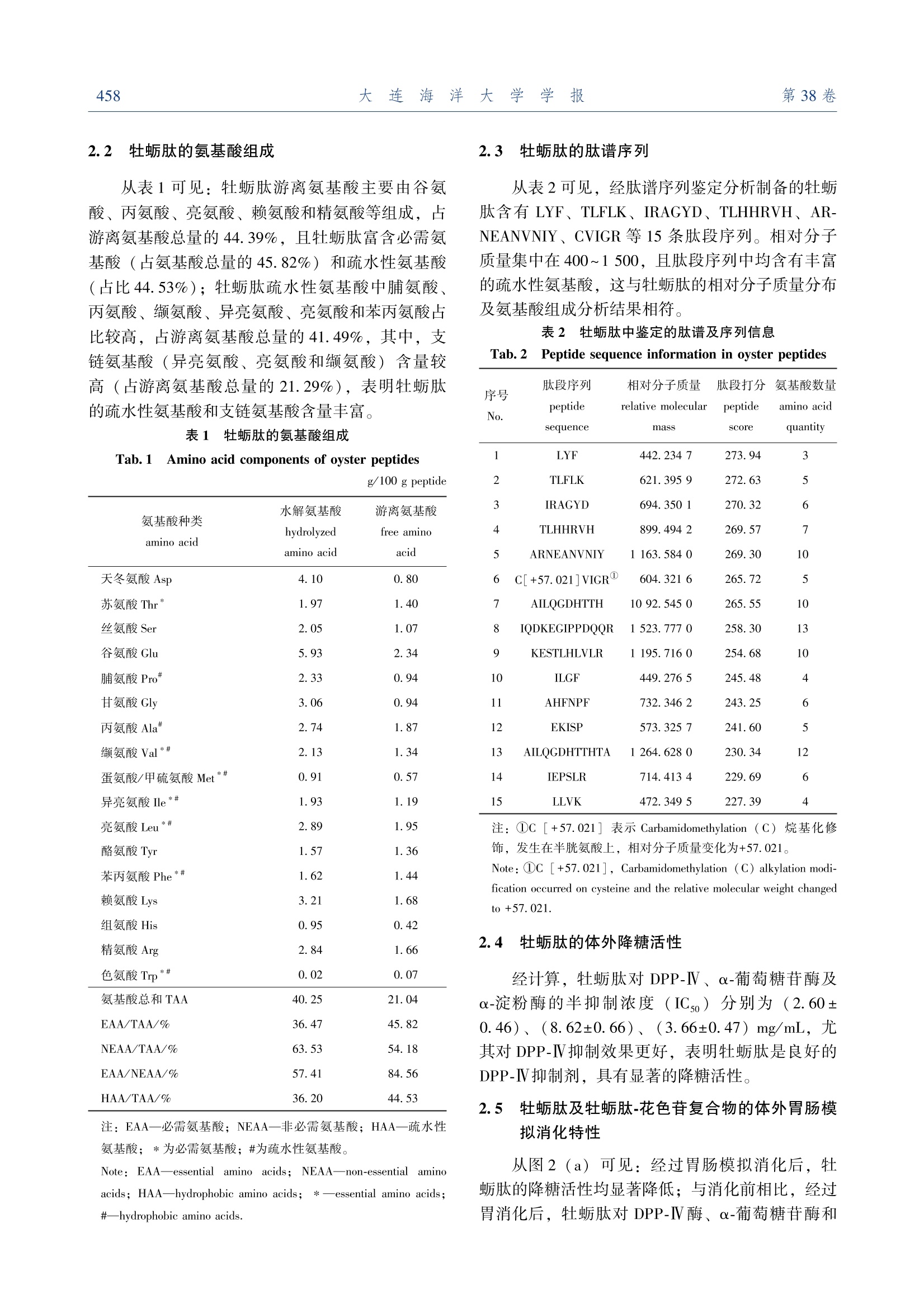
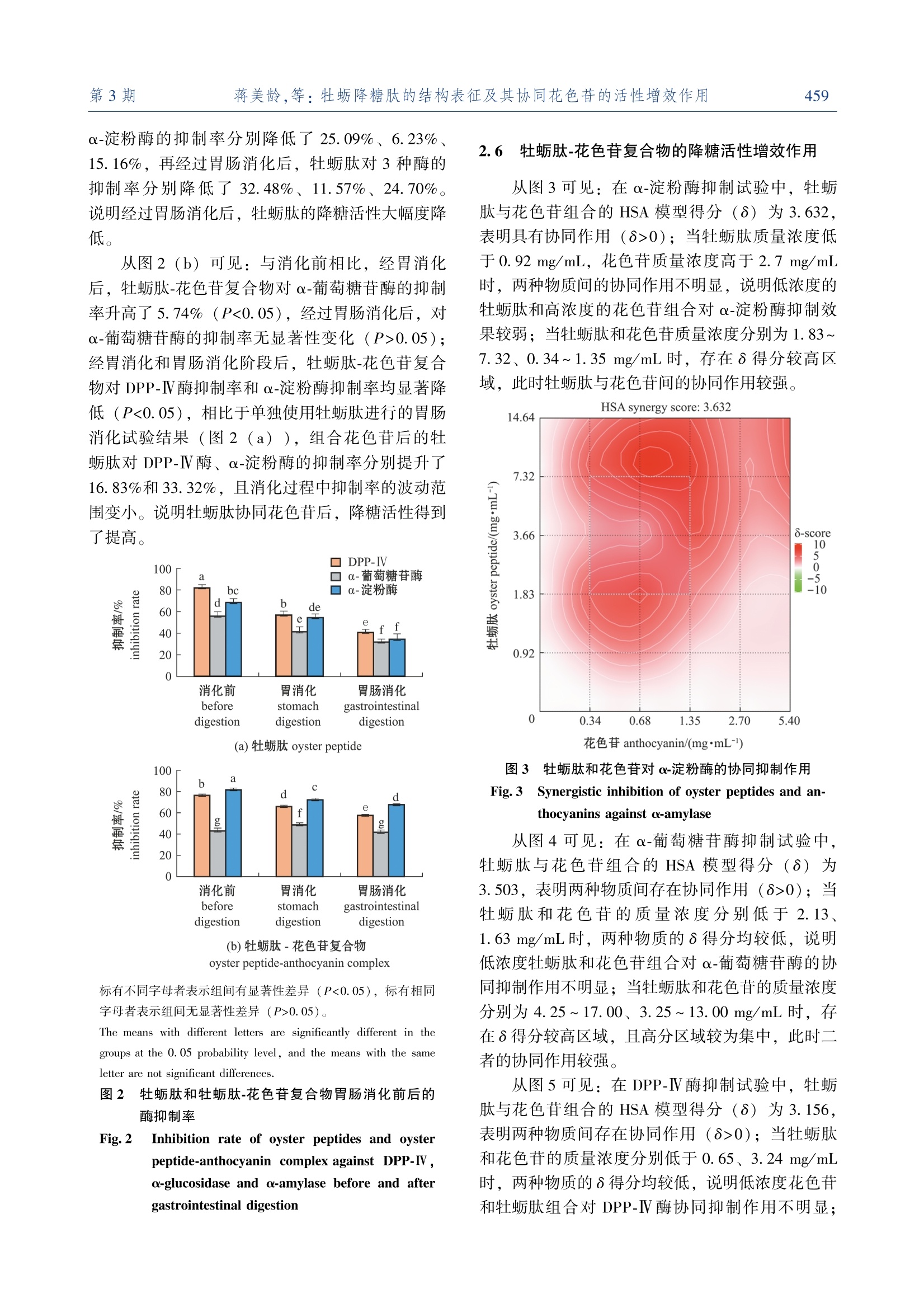
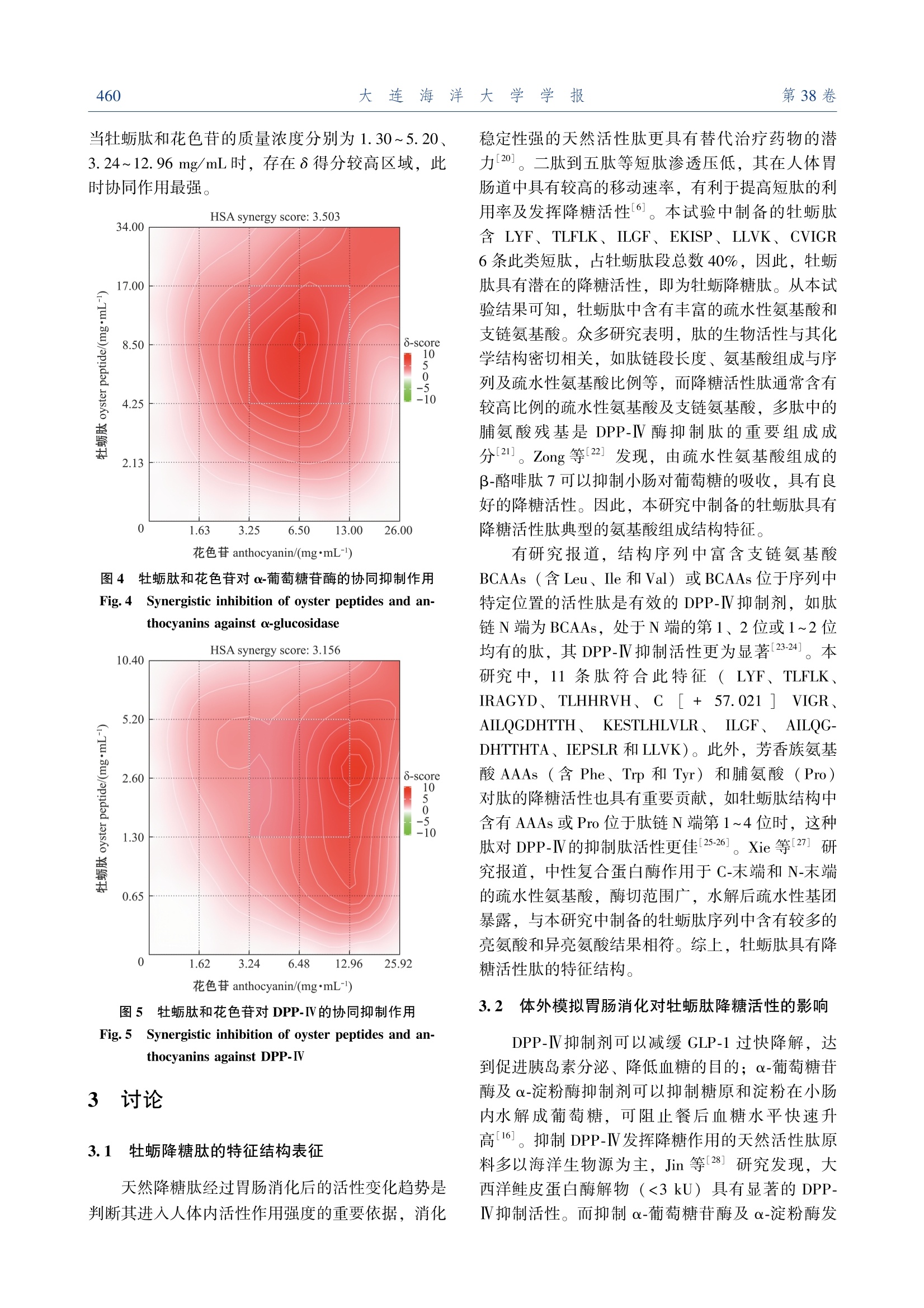
牡蛎降糖肽的结构表征及其协同花色苷的活性增效作用大连海洋大学学报JOURNAL OF DALIAN OCEAN UNIVERSITYVol. 38 No. 3June 2 0 2 3第 38卷第 3期2 0 2 3年 6月文章编号:2095-1388(2023)03-0455-09 大连海洋大学学报第38卷654 牡蛎降糖肽的结构表征及其协同花色苷的活性增效作用 蒋美龄,陈忠琴,秦小明,孙旭佳,曹文红,林海生,高加龙 Structural characterization and the synergistic effects with anthocyanin of oyster hypoglycemic peptides JIANG Meiling,CHEN Zhongqin,QIN Xiaoming,SUN Xujia,CAO Wenhong,LIN Haisheng,GAO Jialong 在线阅读 View online: https://doi.org/10.16535/j.cnki.dlhyxb.2022-282 您可能感兴趣的其他文章 Articles you may be interested in 牡蛎ACE抑制肽对原发性高血压大鼠的降压效果及其性质研究 Properties and antihypertensive effect of ACE inhibitory peptides from oyster on spontaneously hypertensive rats 大连海洋大学学报. 2018, 33(6): 788 https://doi.org/10.16535/j.cnki.dlhyxb.2018.06.017 冰温酶解贮藏对低盐脱水牡蛎滋味的影响 Effects of ice temperature enzymatic hydrolysis on taste of low salt dehydrated oyster during storage 大连海洋大学学报. 2021, 36(5): 815 https://doi.org/10.16535/j.cnki.dlhyxb.2020-317 牡蛎营养特性及功能活性研究进展 Research progress on nutritional characteristics and bioactivities of oysters: a review 大连海洋大学学报. 2022, 37(5): 719 https://doi.org/10.16535/j.cnki.dlhyxb.2022-243 鱼肽和蛋白酶替代部分鱼粉对凡纳滨对虾生长、消化酶及肠道菌群的影响 Effects of partial fish meal substitution by fish peptide and protease on growth, intestinal digestive enzymes and intestinal microflora of Pacific white shrimp Litopenaeus vannamei 大连海洋大学学报. 2022, 37(5): 756 https://doi.org/10.16535/j.cnki.dlhyxb.2021-194 拟穴青蟹抗菌肽SCY2在毕赤酵母中的表达及其抗菌活性 Expression, purification and antimicrobial activity of SCY2 from mud crab Scylla paramamosain in yeast Pichia pastoris 大连海洋大学学报. 2018, 33(2): 151 https://doi.org/10.16535/j.cnki.dlhyxb.2018.02.003 DOI :10. 16535 / j. cnki. dlhyxb. 2022-282 蒋美龄 1,陈忠琴 1,2∗,秦小明 1,3∗,孙旭佳 1,曹文红 1,2,3,林海生 1,3,高加龙 1,3 (1.广东海洋大学食品科技学院 ,国家贝类加工技术研发分中心 (湛江 ),广东省水产品加工与安全重点实验室 ,广东省海洋食 品工程技术研究中心 ,广东省海洋生物制品工程实验室 ,水产品深加工广东普通高等学校重点实验室 ,广东湛江 524088;2.广东海洋大学深圳研究院 ,广东深圳 518108; 3.大连工业大学海洋食品精深加工关键技术省部共建协同创新中心 ,辽宁 大连 116034) 摘要 :为探讨牡蛎肽 (oyster peptide)的降糖作用及消化稳定性 ,以香港牡蛎 (Grassostrea hongkongensis )为原料制备牡蛎肽 ,对其结构特征进行表征 ,通过体外降糖模型及模拟胃肠消化模型考察牡蛎肽的降糖活 性及消化稳定性 ,并利用花色苷协同提高牡蛎肽消化稳定性和降糖活性 。结果表明 :牡蛎肽含有丰富的疏 水性氨基酸 ,相对分子质量主要集中在 1 000之内 ;肽谱分析显示 ,牡蛎肽含有 LYF、 TLFLK、 IRAGYD、TLHHRVH、 ARNEANVNIY、 CVIGR 等 15条肽段 ,具有显著的降糖肽结构特征 ;牡蛎肽对 α-淀粉酶 、 α-葡 萄糖苷酶和二肽基肽酶 Ⅳ (DPP-Ⅳ)的半抑制浓度 (IC50)分别为 (3.66±0.47)、 (8.62±0.66)、 (2.60±0.46)mg/mL,具有良好的降糖活性 ;牡蛎肽的消化稳定性相对较差 ,经体外模拟胃肠消化后 ,其 α-淀粉 酶 、 α-葡萄糖苷酶和 DPP-Ⅳ酶的抑制率显著下降了 6.23%~32.48% (P <0.05);通过组合花色苷设计协同 矩阵 ,选用 Highest Single Agent (HSA)模型计算组合效果 ,结果显示 ,牡蛎肽和花色苷对 α-葡萄糖苷酶 (HSA 协同得分 3.503)、 α-淀粉酶 (HSA 协同得分 3.632)和 DPP-Ⅳ (HSA 协同得分 3.156)具有协同抑 制作用 ,经体外模拟胃肠消化试验验证 ,花色苷可以提高牡蛎肽的消化稳定性 。研究表明 ,牡蛎肽具有降 糖活性肽的特征结构 ,其与花色苷协同后具有显著的消化稳定性和降糖活性增效作用 。 关键词 :牡蛎肽 ;结构表征 ;降糖活性 ;花色苷 ;增效作用 中图分类号 : S 986.2 文献标志码 : A 糖尿病是一种以高血糖为特征的代谢性疾病 ,可诱发心肌梗死 、中风和心血管疾病等各种慢性并 发症 。近年来 ,糖尿病发病率在全世界迅速增加 ,已成为继心脑血管疾病 、癌症后的第三大危害人类 生命健康的疾病 [1]。据统计 , 2021年中国的糖尿 病患者数量为 1.4亿 , 10年间增幅达 56%,预计 2030年中国糖尿病患者人数将超过 1.6亿 [2]。 Ⅱ型糖尿病是糖尿病患者的主要发病类型 ,现有研究 表明 , α-葡萄糖苷酶 、 α-淀粉酶和二肽基肽酶 Ⅳ(DPP-Ⅳ)是治疗 Ⅱ型糖尿病的重要靶点 [3]。 α-葡 萄糖苷酶可直接参与多糖代谢 ,通过水解葡萄糖苷 键在小肠释放葡萄糖 ; α-淀粉酶通过水解淀粉内部 α-1, 4-糖苷键生成葡萄糖 ,促进血糖快速升高 。因此 ,抑制 α-葡萄糖苷酶及 α-淀粉酶的活性可延 缓多糖向单糖的转化 ,维持血糖正常水平 ,并有效 改善餐后高血糖症状 。胰高血糖素样肽 -1 (GLP-1)是一种能刺激胰岛素分泌的激素 ,可系统调控 血糖水平 ,但体内 GLP-1极易被 DPP-IV 降解失 活 ,而 DPP-IV 抑制剂可延长 GLP-1的作用时间 ,达到降低血糖的目的 。因此 , α-淀粉酶 、 α-葡萄糖 苷酶和 DPP-Ⅳ酶天然抑制剂具有调控血糖的良好 潜力 。 收稿日期 : 2022-09-20 基金项目 :国家自然科学基金 ( 32201971 );广东省基础与应用基础研究基金 ( 2021A1515110621 );湛江市科技计划项目 (060302042007);财政部和农业农村部 :国家现代农业产业技术体系资 秦小明 (1964—),男 ,教授 。 E-mail:xiaoming0502@ 21cn. com 牡蛎 ,俗称 “蚝 ”,是世界第一大养殖贝类 ,是中国卫生部门批准的第一批药食同源的海产品之 一 [4]。牡蛎富含优质蛋白质 ,有 “海洋牛奶 ”之 美誉 ,是制备活性肽的良好来源 。研究表明 ,牡蛎 肽 (oyster peptide)具有调节免疫力 、降血压及降 血糖等多种生理活性 [5-6]。牡蛎肽因具备特殊海洋 环境赋予的独特氨基酸序列结构 ,在降糖应用方面 具有天然优势 [7-8]。但其消化稳定性差的问题 ,极 大地限制了活性肽的活性发挥 [9]。花色苷是一类 广泛存在于果蔬中的天然色素 ,具有显著的降血糖 功能 [10-11],且本课题组前期研究发现 ,花色苷与 牡蛎肽的复合物具有提高二者消化稳定性和降糖活 性的潜力 。目前 ,关于牡蛎降糖肽的结构特征 、消 化稳定性及活性增效研究鲜有报道 。本研究中 ,以 香港牡蛎 (Grassostrea hongkongensis )为原料制备 牡蛎降糖肽 ,对其相对分子质量分布 、氨基酸组成 和肽谱序列等结构特征进行表征 ;通过体外降糖模 型 (α-葡萄糖苷酶 、 α-淀粉酶和 DPP-Ⅳ抑制模型 )及模拟胃肠消化模型考察牡蛎肽的降糖活性及消化 稳定性 ,并探究了利用花色苷协同提高牡蛎肽消化 稳定性和降糖活性 ,以期为新型牡蛎降糖肽及其活 性增效研究提供科学参考 。 1 材料与方法 1.1 材料 试验用新鲜牡蛎肉购于广东省湛江市水产品批 发市场 ;黑豆皮花色苷购于西安维珍生物科技有限 公司 。试剂 :动物蛋白水解复合酶 (10万 U/ g)购 于广西南宁庞博生物工程有限公司 ; α-葡萄糖苷酶 (70万 U/mL)、 α-淀粉酶 (猪胰腺 , 5 U/mg)、可 溶性淀粉 、对硝基苯基吡喃葡萄糖苷 (p-NPG)、胃蛋白酶 、胰蛋白酶和磷酸缓冲液 (PBS)均购于 上海源叶生物科技有限公司 ;甲醛 (色谱级 )购 于北京索莱宝科技有限公司 ;氮 -甘氨酰脯氨酰 -对 硝基苯胺盐酸盐 (328.75 mol / g)、二肽基肽酶 Ⅳ(4 000 U/μg)购于美国 Sigma 公司 。 仪器与设备 : Varioskan Flash 全自动酶标仪 (美国 Thermo 公司 ); 5810R 高速台式冷冻离心机 (美国 Eppendorf 公司 );半制备液相色谱仪 (美国 安捷伦公司 ); FD-551大型立式冷冻干燥机 (东京 理化器械公司 );旋蒸浓缩系统 N-4000 (东京理化 器械公司 );全自动凯氏定氮仪 VAPODEST 450(德国格哈特分析仪器有限公司 )。 1.2 方法 1.2. 1 牡蛎肽的制备 参考柏昌旺等 [12]方法优 化牡蛎肽制备条件 。将香港牡蛎肉经流水解冻后按 料液比 1 ∶ 3. 9 (g ∶ mL)加水匀浆 ,加入动物蛋 白酶 (每 g 牡蛎全肉加酶量为 3 300 U)酶解 3 h (pH 6~7, 47 ℃),酶解结束后沸水浴灭酶 10 min,迅速冷却至室温 , 4 ℃下以 8 000r / min 离心 15 min 后 ,用 4层脱脂棉纱布过滤 ,取上清液冷冻 干燥 48 h 备用 。 1.2. 2 牡蛎肽相对分子质量分布 参考 Li 等 [13]的方法 ,分析牡蛎肽的相对分子质量分布 。 样品制备 :称取样品 100 mg 于 10 mL 容量瓶 中 ,用流动相稀释至刻度 ,超声 5 min 后离心 ,采 用孔径为 0.22 μm 的有机系滤膜过滤后进样 ,同 时配制质量浓度为 1 mg/mL 的多肽标准溶液 。 色谱条件 :色谱柱 , TSKgel2000SWXL300 mm×7.8 mm;流动相为乙腈 +水 +三氟乙酸 ,三者的体 积比为 40 ∶ 60 ∶ 0. 1;流速为 0.5 mL/min,柱温为 30 ℃,进样体积为 20 μL,检测波长为 220 nm。对多肽标准品的保留时间及相对分子质量的对数进 行线性拟合 ,制作标准曲线 ,并采用面积归一化法 进行相对分子质量分布的计算 。 1.2. 3 牡蛎肽氨基酸组成分析 按照 GB 5009.124—2016 《食品安全国家标准食品中氨基酸 的测定 》中的方法 ,采用全自动氨基酸分析仪进 行氨基酸组成分析 。 1.2. 4 牡蛎肽肽谱序列测定 参考 Xu 等 [14]的方 法并稍作改进 。采用 UPLC-Q-TOF 技术对牡蛎肽进 行肽谱序列鉴定 。选取 C18色谱柱 (150 mm ×2.0 mm),检测波长为 220 nm,柱温为 40 ℃,流 速为 0.3 mL/min,进样量为 10 μL。流动相 : A 液 为纯乙腈和 0.1%甲酸 ; B 液为超纯水和 0.1%甲 酸 。采取梯度洗脱方式 : 0 min 时 A 液为 96% (均 为体积分数 ,下同 ),在 55 min 内 B 液由 4%增加 至 40%; 56 min 时 B 液为 95%,并保持至 66 min,结束后用 95%的 B 液冲洗柱子 。 1.2. 5 牡蛎肽体外降糖活性测定 1) α-葡萄糖苷酶抑制率 。参照 De Matos 等 [15]的方法并稍作调整 ,采用 pNPG 法测定牡蛎 肽的 α-葡萄糖苷酶抑制活性 。移取 110 μL 磷酸盐 缓冲液 (pH 6.8)于 96孔板微孔 ,分别加入 20 μL 样品溶液和 20 μL 1 U/mL 的 α-葡萄糖苷酶溶液 ,混匀后于 37 ℃下孵育 10 min 后 ,加入 20 μL 1.25 mmol / L 的底物 pNPG,体系于 37 ℃下反应 20 min 后 ,加入 80 μL 0.1 mol / L 的碳酸钠终止反 应 ,以阿卡波糖作为阳性对照组 ,重复以上试验操 作 。用酶标仪在 405 nm 处测定其吸光值 ,并计算 α-葡萄糖苷酶抑制率 (R Ⅰ),即 式中 : A 样品组 、样品空白 1、 A 2、 A 和 A 分别为 34组 、对照组和空白组的吸光值 。 2) α-淀粉酶抑制率 。参照 De Matos 等 [15]的 方法并稍作调整 ,采用 DNS 比色法测定牡蛎肽的 α-淀粉酶抑制活性 。在玻璃试管中加入 300 μL 样 液和 300 μL 淀粉酶溶液 (0.1 U/mL), 37 ℃下水 浴 15 min 后 ,加入 300 μL 可溶性淀粉 (体积分数 为 1%), 37 ℃下反应 30 min 后 ,加入 500 μL DNS 在沸水浴中继续反应 10 min,冰浴冷却后加入 8 600 μL 蒸馏水 ,漩涡混匀 。以阿卡波糖作为阳 性对照组 ,重复以上试验操作 ,取 200 μL 反应液 置于 96孔板 ,检测其在 540 nm 处的吸光值 ,并根 据式 (1)计算 α-淀粉酶抑制率 。 3) DPP-Ⅳ抑制率 。参考 Lourdes 等 [16]的方 法 ,采用 Gly-Pro 法测定牡蛎肽的 DPP-Ⅳ抑制活 性 。在 96孔板微孔中 ,分别加入 25 μL 样品溶液 和 25 μL 底物 (甘氨酸对硝基苯胺溶液 ),混匀后 于 37 ℃下孵育 10 min 后 ,加入 50 μL 0.01 U/mL 的 DPP-Ⅳ,体系于 37 ℃下反应 60 min 后 ,加入 100 μL 1 mol / L 的醋酸钠缓冲液 (pH 4)终止反 应 。以西格列汀作为阳性对照组 ,重复以上试验操 作 。采用酶标仪在 405 nm 处测定其吸光值 ,并计 算 DPP-Ⅳ抑制率 ,即 式中 : A 1和 A 2分别为对照组和样品组的吸光值 。 1.2. 6 牡蛎肽 -花色苷复合物的体外模拟胃肠消化 特性分析 参照 Guo 等 [17]的方法并稍作调整 。称 取 1.0 g 牡蛎肽放入 100 mL 烧杯中 ,并加入 0.2 g NaCl 和 50 mL 去离子水 ,用浓度为 1.0 mol / L 的 HCl 溶液调节 pH 至 2.0,加入 0.05 g 胃蛋白酶后 于 37 ℃水浴反应 2 h,消化结束后用 100 ℃沸水灭 酶 ,冷却至室温后补水至 100 mL。移取 50 mL 胃 消化液用水定容至 100 mL,牡蛎肽在反应体系的 质量浓度为 5 mg/mL,将稀释后的胃消化液于 4 ℃下以 8 000 r / min 离心 15 min,取上清液进行 α-葡 萄糖苷酶 、 α-淀粉酶和 DPP-Ⅳ抑制活性测定 。 按上述方法进行胃肠消化试验 ,在上述剩余的 50 mL 胃消化液中继续加入 2 mL 胰蛋白酶溶液 , 改用 1.0 mol / L NaOH 溶液调节 pH 至 7.0,其他操 作不变 ,测定胃肠消化液对 3种酶的抑制活性 。 重复上述试验步骤 ,测定经胃消化及胃肠消化 后牡蛎肽 -花色苷复合物的体外降糖活性 ,考察牡 蛎肽 -花色苷复合物的消化稳定性 。 1.2. 7 牡蛎肽 -花色苷复合物的降糖活性增效作用 试验 参考 Aleksandr 等 [18]的方法 ,设计牡蛎肽 与花色苷的样品剂量协同矩阵 ,根据矩阵对应的不 同样品组合浓度进行 α-葡萄糖苷酶 、 α-淀粉酶和 DPP-Ⅳ抑制活性测定 ,利用多药物联用效应计算 软件 SynergyFinder ( https:/ / synergyfinder. org / )对所得的试验数据进行分析 。该软件中的 HSA 模 型在评价协同效果试验中运用广泛 ,采用 HSA 模 型计算得到 Synergyscore ( δ ),当 δ =0、δ >0和 δ <0时 ,分别对应于样品组合间的零交互 、协同和 拮抗作用 [19]。 1.3 数据处理 试验数据均采用平均值 ±标准差 (mean ±S. D. )表示 ,每组试验均设置 3个平行 。采用 SPSS 22.0软件对试验数据进行单因素方差分析 ,采用 Duncan 法进行组间多重比较 ,显著性水平设 为 0.05。 2 结果与分析 2.1 牡蛎肽的相对分子质量分布 由中性复合蛋白酶酶解得到的牡蛎肽经色谱柱 洗脱后 ,出现 4个明显的吸收峰 。结合峰面积归一 化法分析显示 ,牡蛎肽主要是由相对分子质量为 1 000以下的小分子多肽组成 ,占比高达 84.01%,其中 ,相对分子质量为 180~500的短肽占比达 46.70% (图 1),说明制备的牡蛎肽为低聚肽 ,且 主要是二肽到五肽之间的小分子肽 。 相对分子质量 relative molecul a r mass 图 1 牡蛎肽的相对分子质量分布 Fig.1 Relative molecular mass distribution of oyster peptides 2.2 牡蛎肽的氨基酸组成 从表 1可见 :牡蛎肽游离氨基酸主要由谷氨 酸 、丙氨酸 、亮氨酸 、赖氨酸和精氨酸等组成 ,占 游离氨基酸总量的 44.39%,且牡蛎肽富含必需氨 基酸 (占氨基酸总量的 45.82%)和疏水性氨基酸 (占比 44.53%);牡蛎肽疏水性氨基酸中脯氨酸 、丙氨酸 、缬氨酸 、异亮氨酸 、亮氨酸和苯丙氨酸占 比较高 ,占游离氨基酸总量的 41.49%,其中 ,支 链氨基酸 (异亮氨酸 、亮氨酸和缬氨酸 )含量较 高 (占游离氨基酸总量的 21.29%),表明牡蛎肽 的疏水性氨基酸和支链氨基酸含量丰富 。 表 1 牡蛎肽的氨基酸组成 Tab.1 Amino acid components of oyster peptides g / 100 g peptide 氨基酸种类 amino acid 水解氨基酸 hydrolyzed amino acid 游离氨基酸 free amino acid 天冬氨酸Asp 4. 10 0. 80 苏氨酸 Thr∗ 1. 97 1. 40 丝氨酸 Ser 2. 05 1. 07 谷氨酸Glu 5. 93 2. 34 脯氨酸 Pro# 2. 33 0. 94 甘氨酸Gly 3. 06 0. 94 丙氨酸Ala# 2. 74 1. 87 缬氨酸Val∗# 2. 13 1. 34 蛋氨酸 /甲硫氨酸Met∗# 0. 91 0. 57 异亮氨酸 Ile∗# 1. 93 1. 19 亮氨酸 Leu∗# 2. 89 1. 95 酪氨酸 Tyr 1. 57 1. 36 苯丙氨酸 Phe∗# 1. 62 1. 44 赖氨酸 Lys 3. 21 1. 68 组氨酸His 0. 95 0. 42 精氨酸Arg 2. 84 1. 66 色氨酸 Trp∗# 0. 02 0. 07 氨基酸总和 TAA 40. 25 21. 04 EAA/ TAA/% 36. 47 45. 82 NEAA/ TAA/% 63. 53 54. 18 EAA/NEAA/% 57. 41 84. 56 HAA/ TAA/% 36. 20 44. 53 注 : EAA—必需氨基酸 ; NEAA—非必需氨基酸 ; HAA—疏水性 氨基酸 ; ∗为必需氨基酸 ;#为疏水性氨基酸 。 Note: EAA—essential amino acids; NEAA—non-essential amino acids; HAA—hydrophobic amino acids; ∗—essential amino acids;#—hydrophobic amino acids. 2.3 牡蛎肽的肽谱序列 从表 2可见 ,经肽谱序列鉴定分析制备的牡蛎 肽含有 LYF、 TLFLK、 IRAGYD、 TLHHRVH、 AR-NEANVNIY、 CVIGR 等 15条肽段序列 。相对分子 质量集中在 400~1 500,且肽段序列中均含有丰富 的疏水性氨基酸 ,这与牡蛎肽的相对分子质量分布 及氨基酸组成分析结果相符 。 表 2 牡蛎肽中鉴定的肽谱及序列信息 Tab.2 Peptide sequence information in oyster peptides 序号 No. 肽段序列 peptide sequence 相对分子质量 relative molecular mass 肽段打分 peptide score 氨基酸数量 amino acid quantity 1 LYF 442. 234 7 273. 94 3 2 TLFLK 621. 395 9 272. 63 5 3 IRAGYD 694. 350 1 270. 32 6 4 TLHHRVH 899. 494 2 269. 57 7 5 ARNEANVNIY 1 163. 584 0 269. 30 10 6 C[+57. 021]VIGR① 604. 321 6 265. 72 5 7 AILQGDHTTH 10 92. 545 0 265. 55 10 8 IQDKEGIPPDQQR 1 523. 777 0 258. 30 13 9 KESTLHLVLR 1 195. 716 0 254. 68 10 10 ILGF 449. 276 5 245. 48 4 11 AHFNPF 732. 346 2 243. 25 6 12 EKISP 573. 325 7 241. 60 5 13 AILQGDHTTHTA 1 264. 628 0 230. 34 12 14 IEPSLR 714. 413 4 229. 69 6 15 LLVK 472. 349 5 227. 39 4 注 : ①C [+57. 021]表示 Carbamidomethylation(C)烷基化修 饰 ,发生在半胱氨酸上 ,相对分子质量变化为 +57. 021。 Note: ①C [+57. 021], Carbamidomethylation(C)alkylation modi-fication occurred on cysteine and the relative molecular weight changed to +57. 021. 2.4 牡蛎肽的体外降糖活性 经计算 ,牡蛎肽对 DPP-Ⅳ、 α-葡萄糖苷酶及 α-淀粉酶的半抑制浓度 (IC50)分别为 (2.60±0.46)、 (8.62±0.66)、 (3.66±0.47)mg/mL,尤 其对 DPP-Ⅳ抑制效果更好 ,表明牡蛎肽是良好的 DPP-Ⅳ抑制剂 ,具有显著的降糖活性 。 2.5 牡蛎肽及牡蛎肽 -花色苷复合物的体外胃肠模 拟消化特性 从图 2 (a)可见 :经过胃肠模拟消化后 ,牡 蛎肽的降糖活性均显著降低 ;与消化前相比 ,经过 胃消化后 ,牡蛎肽对 DPP-Ⅳ酶 、 α-葡萄糖苷酶和 α-淀粉酶的抑制率分别降低了 25.09%、 6.23%、15.16%,再经过胃肠消化后 ,牡蛎肽对 3种酶的 抑制率分别降低了 32.48%、 11.57%、 24.70%。说明经过胃肠消化后 ,牡蛎肽的降糖活性大幅度降 低 。从图 2 (b)可见 :与消化前相比 ,经胃消化 后 ,牡蛎肽 -花色苷复合物对 α-葡萄糖苷酶的抑制 率升高了 5.74% (P <0.05),经过胃肠消化后 ,对 α-葡萄糖苷酶的抑制率无显著性变化 (P >0.05);经胃消化和胃肠消化阶段后 ,牡蛎肽 -花色苷复合 物对 DPP-Ⅳ酶抑制率和 α-淀粉酶抑制率均显著降 低 (P <0.05),相比于单独使用牡蛎肽进行的胃肠 消化试验结果 (图 2 (a) ),组合花色苷后的牡 蛎肽对 DPP-Ⅳ酶 、 α-淀粉酶的抑制率分别提升了 16.83%和 33.32%,且消化过程中抑制率的波动范 围变小 。说明牡蛎肽协同花色苷后 ,降糖活性得到 了提高 。 标有不同字母者表示组间有显著性差异 (P <0. 05),标有相同 字母者表示组间无显著性差异 (P >0. 05)。 The means withdifferent letters are significantly different in the groups at the 0. 05 probability level, and the means with the same letter are not significant differences. 图 2 牡蛎肽和牡蛎肽 -花色苷复合物胃肠消化前后的 酶抑制率 Fig. 2 Inhibition rate of oyster peptides and oyster peptide-anthocyanin complex against DPP-Ⅳ,α-glucosidase and α-amylase before and after gastrointestinal digestion 2.6 牡蛎肽 -花色苷复合物的降糖活性增效作用 从图 3可见 :在 α-淀粉酶抑制试验中 ,牡蛎 肽与花色苷组合的 HSA 模型得分 ( δ )为 3.632,表明具有协同作用 ( δ >0);当牡蛎肽质量浓度低 于 0.92 mg/mL,花色苷质量浓度高于 2.7 mg/mL 时 ,两种物质间的协同作用不明显 ,说明低浓度的 牡蛎肽和高浓度的花色苷组合对 α-淀粉酶抑制效 果较弱 ;当牡蛎肽和花色苷质量浓度分别为 1.83~7.32、 0.34~1.35 mg/mL 时 ,存在 δ 得分较高区 域 ,此时牡蛎肽与花色苷间的协同作用较强 。 花色苷 a nthoc yan in/(mg*mL -) 图 3 牡蛎肽和花色苷对 α-淀粉酶的协同抑制作用 Fig. 3 Synergistic inhibition of oyster peptides and an-thocyanins against α-amylase 从图 4可见 :在 α-葡萄糖苷酶抑制试验中 ,牡蛎肽与花色苷组合的 HSA 模型得分 ( δ )为 3.503,表明两种物质间存在协同作用 ( δ >0);当 牡蛎肽和花色苷的质量浓度分别低于 2.13、1.63 mg/mL 时 ,两种物质的 δ 得分均较低 ,说明 低浓度牡蛎肽和花色苷组合对 α-葡萄糖苷酶的协 同抑制作用不明显 ;当牡蛎肽和花色苷的质量浓度 分别为 4.25~17.00、 3.25~13.00 mg/mL 时 ,存 在 δ 得分较高区域 ,且高分区域较为集中 ,此时二 者的协同作用较强 。 从图 5可见 :在 DPP-Ⅳ酶抑制试验中 ,牡蛎 肽与花色苷组合的 HSA 模型得分 ( δ )为 3.156,表明两种物质间存在协同作用 ( δ >0);当牡蛎肽 和花色苷的质量浓度分别低于 0.65、 3.24 mg/mL 时 ,两种物质的 δ 得分均较低 ,说明低浓度花色苷 和牡蛎肽组合对 DPP-Ⅳ酶协同抑制作用不明显 ; 当牡蛎肽和花色苷的质量浓度分别为 1.30~5.20、3.24~12.96 mg/mL 时 ,存在 δ 得分较高区域 ,此 时协同作用最强 。 花色苷 a n t h o cy an i n /(m g *m L -) 图 4 牡蛎肽和花色苷对 α-葡萄糖苷酶的协同抑制作用 Fig. 4 Synergistic inhibition of oyster peptides and an-thocyanins against α-glucosidase 花色苷 an tho cya n i n /(mg*mL -) 图 5 牡蛎肽和花色苷对 DPP-Ⅳ的协同抑制作用 Fig. 5 Synergistic inhibition of oyster peptides and an-thocyanins against DPP-Ⅳ 3 讨论 3.1 牡蛎降糖肽的特征结构表征 天然降糖肽经过胃肠消化后的活性变化趋势是 判断其进入人体内活性作用强度的重要依据 ,消化 稳定性强的天然活性肽更具有替代治疗药物的潜 力 [20]。二肽到五肽等短肽渗透压低 ,其在人体胃 肠道中具有较高的移动速率 ,有利于提高短肽的利 用率及发挥降糖活性 [6]。本试验中制备的牡蛎肽 含 LYF、 TLFLK、 ILGF、 EKISP、 LLVK、 CVIGR 6条此类短肽 ,占牡蛎肽段总数 40%,因此 ,牡蛎 肽具有潜在的降糖活性 ,即为牡蛎降糖肽 。从本试 验结果可知 ,牡蛎肽中含有丰富的疏水性氨基酸和 支链氨基酸 。众多研究表明 ,肽的生物活性与其化 学结构密切相关 ,如肽链段长度 、氨基酸组成与序 列及疏水性氨基酸比例等 ,而降糖活性肽通常含有 较高比例的疏水性氨基酸及支链氨基酸 ,多肽中的 脯氨酸残基是 DPP-Ⅳ酶抑制肽的重要组成成 分 [21]。 Zong 等 [22]发现 ,由疏水性氨基酸组成的 β-酪啡肽 7可以抑制小肠对葡萄糖的吸收 ,具有良 好的降糖活性 。因此 ,本研究中制备的牡蛎肽具有 降糖活性肽典型的氨基酸组成结构特征 。 有研究报道 ,结构序列中富含支链氨基酸 BCAAs (含 Leu、 Ile 和 Val)或 BCAAs 位于序列中 特定位置的活性肽是有效的 DPP-Ⅳ抑制剂 ,如肽 链 N 端为 BCAAs,处于 N 端的第 1、 2位或 1~2位 均有的肽 ,其 DPP-Ⅳ抑制活性更为显著 [23-24]。本 研究中 , 11条肽符合此特征 (LYF、 TLFLK、IRAGYD、 TLHHRVH、 C [ + 57.021 ] VIGR、AILQGDHTTH、 KESTLHLVLR、 ILGF、 AILQG-DHTTHTA、 IEPSLR 和 LLVK)。此外 ,芳香族氨基 酸 AAAs (含 Phe、 Trp 和 Tyr)和脯氨酸 (Pro)对肽的降糖活性也具有重要贡献 ,如牡蛎肽结构中 含有 AAAs 或 Pro 位于肽链 N 端第 1~4位时 ,这种 肽对 DPP-Ⅳ的抑制肽活性更佳 [25-26]。 Xie 等 [27]研 究报道 ,中性复合蛋白酶作用于 C-末端和 N-末端 的疏水性氨基酸 ,酶切范围广 ,水解后疏水性基团 暴露 ,与本研究中制备的牡蛎肽序列中含有较多的 亮氨酸和异亮氨酸结果相符 。综上 ,牡蛎肽具有降 糖活性肽的特征结构 。 3.2 体外模拟胃肠消化对牡蛎肽降糖活性的影响 DPP-Ⅳ抑制剂可以减缓 GLP-1过快降解 ,达 到促进胰岛素分泌 、降低血糖的目的 ; α-葡萄糖苷 酶及 α-淀粉酶抑制剂可以抑制糖原和淀粉在小肠 内水解成葡萄糖 ,可阻止餐后血糖水平快速升 高 [16]。抑制 DPP-Ⅳ发挥降糖作用的天然活性肽原 料多以海洋生物源为主 , Jin 等 [28]研究发现 ,大 西洋鲑皮蛋白酶解物 (<3 kU)具有显著的 DPP-Ⅳ抑制活性 。而抑制 α-葡萄糖苷酶及 α-淀粉酶发 挥降糖作用的天然活性肽原料多以植物源为主 。李 艳敏等 [29]研究发现 ,裙带菜酶解物对 α-葡萄糖苷 酶的 IC50值为 46.08 mg/mL。袁娟等 [30]研究发现 ,白豆提取物对 α-淀粉酶的 IC50值为 4.20 mg/mL。本研究中 ,牡蛎肽对 DPP-Ⅳ、 α-葡萄糖苷酶和 α-淀粉酶的 IC50值均小于 10 mg/mL,因此 ,牡蛎肽 对以上 3种酶均具有良好的抑制作用 ,在作为天然 降糖活性物质上具有良好的应用潜力 ,且作为低分 子量的天然活性肽 ,牡蛎肽具备吸收快 、活性强的 特点 ,符合未来食品行业天然降糖制剂的发展趋 势 。本研究中 ,通过测定胃肠消化前后牡蛎肽对 DPP-Ⅳ、 α-葡萄糖苷酶和 α-淀粉酶的抑制率 ,以 评价降糖活性变化 。结果表明 ,经过胃肠模拟消化 后牡蛎肽对 3种酶的抑制率降低幅度较大 (最高 达 32.48%),可能是由于胃液中的强酸性条件 、胃肠间的 pH 急剧变化 ,以及胃肠液中的消化酶破 坏了牡蛎降糖肽的结构 ,即牡蛎降糖肽的消化稳定 性较差 ,但消化后其仍具有一定的降糖活性 ,因 此 ,有必要采取一定手段提升牡蛎肽的消化稳定 性 ,保留其降糖活性 。 3.3 协同花色苷对牡蛎肽降糖活性的影响 牡蛎肽的消化稳定性较差 ,经过胃肠消化后的 降糖活性损失较大 ,因此 ,考虑通过牡蛎肽协同花 色苷来提高牡蛎肽的消化稳定性 ,提高其降糖活 性 。本研究中 ,牡蛎肽 -花色苷复合物在体外胃肠 模拟消化过程中 ,对 DPP-Ⅳ、 α-葡萄糖苷酶和 α-淀粉酶的抑制率变化范围较小 ,与牡蛎肽相比 ,模 拟消化后对 DPP-Ⅳ酶 、 α-淀粉酶的抑制率有显著 提高 ,这可能是复合物间形成了较稳定的体系 ,可 对抗复杂的模拟消化环境 ,保护了具有降糖活性的 物质 。而对 α-葡萄糖苷酶的抑制率呈先上升后降 低的趋势 ,这可能是花色苷在酸性环境的胃消化阶 段较稳定 ,发挥了最大的活性作用 ,在回到中性环 境的肠消化阶段后 ,对 α-葡萄糖苷酶的抑制作用 又回到消化前水平 。因此 ,牡蛎肽协同花色苷后可 提高消化稳定性和降糖活性 。 为进一步研究牡蛎肽 -花色苷复合物的降糖活 性增效作用 ,采用协同作用研究软件 Synergyfinder 中的 HSA 模型算法 ,评价牡蛎肽与花色苷对 α-淀 粉酶 、 α-葡萄糖酶及 DPP-Ⅳ的协同抑制作用 。结 果表明 ,牡蛎肽 -花色苷复合物对 α-淀粉酶 、 α-葡 萄糖酶及 DPP-Ⅳ抑制试验的 HSA 模型得分 ( δ )均大于 0,说明牡蛎肽与花色苷对这 3种酶具有协 同抑制作用 。这可能是复合物中花色苷的羟基与牡 蛎肽的氨基酸残基发生了相互作用 ,对二者的降糖 活性结构产生了保护作用 ,提高了复合物对这 3种 酶的抑制率 ,使复合物具有协同降糖作用 。 4 结论 1)制备的牡蛎肽相对分子质量主要集中在 1 000以下 ,其必需氨基酸和疏水性氨基酸含量丰 富 。肽谱序列鉴定发现 ,牡蛎降糖肽富含支链氨基 酸和脯氨酸 ,且位于肽链 N 端 1~4位的特定位置 ,说明牡蛎肽具备典型的降糖肽特征结构 。 2)牡蛎肽对 α-葡萄糖苷酶 、 α-淀粉酶和 DPP-Ⅳ具有显著的抑制效果 ,是良好的降糖肽 。 3)牡蛎肽经过体外模拟胃肠消化后 ,其降糖 活性显著降低 ,说明牡蛎肽消化稳定性差 。通过组 合花色苷设计协同矩阵试验 (以 HSA 为计算模型 )发现 ,牡蛎肽 -花色苷对上述 3种酶具有一定的协 同抑制效果 ,并用复合物再次进行体外模拟胃肠消 化试验 ,确定了花色苷具有协同提高牡蛎肽消化稳 定性的作用 。本研究结果为进一步开展牡蛎降糖肽 的稳态化及活性增效研究提供科学依据 。 参考文献 : [1] 董宇婷 ,王荣春 .降糖肽的发展现状及研究进展 [J].生物信息 学 ,2018,16(2):83-89. DONG Y T,WANG R C. The status quo and research progress of hypoglycemic peptide [ J ]. Chinese Journal of Bioinformatics,2018,16(2):83-89. (in Chinese) [2] 尤莉莉 ,赵金红 ,陈新月 ,等 .国家基本公共卫生服务项目十年 评价 (2009—2019年 )系列报告 (二 ):国家基本公共卫生服务 项目实施十年的进展与成效 [ J].中国全科医学 ,2022(26):3209-3220. YOU L L,ZHAO J H,CHEN X Y,et al National essential public health services programs over the past decade research report two:progress and achievements of the implementation of national essen-tial public health services programs over the past decade[J]. Chi-nese General Practice,2022(26):3209-3220. (in Chinese) [3] ADEFEGHA S A,MOLEHIN O R. Inhibitory activities of brim-stone (Morinda lucida )Roots extract on α-amylase and α-glucosi-dase-In vitro [ J]. Vegetos:An International Journal of Plant Re-search,2017,30(4):105. [4] 蔡水淋 ,陈晓婷 ,刘淑集 ,等 .牡蛎肉的综合开发研究进展 [J].渔业研究 ,2020,42(5):527-532. CAI S L,CHEN X T,LIU S J,et al Research progress in the com-prehensive development of oysters meat [ J]. Journal of Fisheries Research,2020,42(5):527-532. (in Chinese) [5] 蔡亚伟 ,王海燕 ,孟辰 ,等 .牡蛎肉提取物对小鼠抗疲劳和耐缺 氧能力的影响 [ J].山东中医药大学学报 ,2019,43(5):503-507. CAI Y W,WANG H Y,MENG C,et al Effects of oyster meat ex-tract on anti-fatigue ability and hypoxia tolerance in mice[J]. Jour-nal of Shandong University of Traditional Chinese Medicine,2019,43(5):503-507. (in Chinese) [6] 张可佳 ,张胜男 ,祁艳霞 ,等 .牡蛎 ACE 抑制肽对原发性高血压 大鼠的降压效果及其性质研究 [J].大连海洋大学学报 ,2018, 33(6):788-794. ZHANG K J,ZHANG S N,QI Y X,et al. Properties and antihy-pertensive effect of ACE inhibitory peptides from oyster on sponta-neously hypertensive rats[J]. Journal of Dalian Ocean University,2018,33(6):788-794. (in Chinese) [7] XIA E Q,ZHU S S,HE M J,et al Marine peptides as potential a-gents for the management of type 2 diabetes mellitus-A prospect [J] Marine Drugs,2017,15(4):88-103. [8] HAO L L,WANG X C,CAO Y R,et al A comprehensive review of oyster peptides:preparation,characterisation and bioactivities[ J]. Reviews in Aquaculture,2022,14(1):120-138. [9] BECHAUX J,GATELLIER P,LE PAGE J F,et al. A comprehen-sive review of bioactive peptides obtained from animal byproducts and their applications[J]. Food & Function,2019,10(10):6244-6266. [10] MOJICA L,BERHOW M,DE MEJIA E G Black bean anthocya-nin-rich extracts as food colorants:physicochemical stability and antidiabetes potential[J]. Food Chemistry,2017,229:628-639. [11] CHEN Z,WANG C,PAN Y,et al. Hypoglycemic and hypolipi-demic effects of anthocyanins extract from black soybean seed coat in highfatdietandstreptozotocin-induceddiabetic mice [ J]. Food Funct,2018,9(1):426-439. [12] 柏昌旺 ,章超桦 ,林海生 ,等 .响应面法优化制备牡蛎短肽工 艺 [J].广东海洋大学学报 ,2019,39(6):85-92. BAI C W,ZHANG C H,LIN H S,et al. Optimization of prepara-tion process of oyster oligopeptides by response surface methodolo-gy[J]. Journal of Guangdong Ocean University,2019,39(6):85-92. (in Chinese) [13] LI W,XU C,ZHANG C,et al. The purification and identification of immunoregulatory peptides from oyster (Crassostrea hongkon-gensis )enzymatic hydrolysate[J] RSC Advances,2019,9(56):32854-32863. [14] XU L L,HU PP,KONG X L,et al. 3-hydroxypyridinone-l-phen-ylalanine conjugates with antimicrobial and tyrosinase inhibitory activities as potential shrimp preservatives[J]. International Jour-nal of Food Science & Technology,2014,49(3):797-803. [15] DE MATOS F M,GOMES D L J T J,GIOVANNA Z,et al. Pro-duction of black cricket protein hydrolysates with α-amylase,α-glucosidase and angiotensin I-converting enzyme inhibitory activi-ties using a mixture of proteases[J] Biocatalysis and Agricultural Biotechnology,2022,39:102276. [16] LOURDES R F, CAROLINE R, CHEN L. Sequential alcalase and flavourzyme treatment for preparation of α-amylase,α-gluco-sidase,anddipeptidyl peptidase(DPP)-Ⅳinhibitory peptides fromoat protein [ J]:Journal of Functional Foods, 2021, 87:104829. [17] GUO Y X, CHEN X F, GONG P, et al Effects of simulated sali- va-gastrointestinal digestion on the physicochemical properties and bioactivities of Siraitiagrosvenorii polysaccharides [ J]. Interna-tional Journalof FoodScience &Technology, 2022,57 ( 7):4495-4506. [18] ALEKSANDR I,LIYE H,TERO A,et al. SynergyFinder:a web application for analyzing drug combination dose-response matrix data[J] Bioinformatics,2017,33(15):2413-2415. [19] DUARTE D,FALCÃO S I,EL MEHDI I,et al. Honeybee venom synergistically enhances the cytotoxic effect of CNS drugs in HT-29 colon and MCF-7 breast cancer cell lines[J]. Pharmaceutics,2022,14(3):511. [20] GALLEGO M,GROOTAERT C,MORA L,et al. Transepithelial transport of dry-cured ham peptides with ACE inhibitory activity through a Caco-2 cell monolayer[J]. Journal of Functional Foods,2016,21:388-395. [21] YE X Y,XIONG L,FU Q Fet al. Chemical characterization and DPP-Ⅳ inhibitory activity evaluation of tripeptides from Gynura divaricata (L) )DC[J]. Journal of Ethnopharmacology,2022,292:115203. [22] ZONG Y F,CHEN W H,ZHANG Y S,et al Effects of intra-gas-tric beta-casomorphin-7 on somatostatin and gastrin gene expres-sion in rat gastric mucosa[J] World Journal of Gastroenterology,2007,13(14):2094-2099. [23] NONGONIERMA A B,PAOLELLA S MUDGIL P,et al. Identifi-cation of novel dipeptidyl peptidase Ⅳ (DPP-Ⅳ)inhibitory pep-tides incamel milkproteinhydrolysates [ J]. Food Chemistry,2018,244:340-348. [24] WANG Y C,SONG X, FENG Y,et al. Changes in peptidomes and Fischer ratios of corn-derived oligopeptides depending on en-zymehydrolysis approaches [ J ]:Food Chemistry, 2019, 297:124931. [25] GONZÁLEZ-MONTOYA M,HERNÁNDEZ-LEDESMA B,MO-RA-ESCOBEDO R,et al Bioactive peptides from germinated soy-bean with anti-diabetic potential by inhibition of dipeptidyl pepti-dase-Ⅳ,α-amylase,and α-glucosidase enzymes[J]. International Journal of Molecular Sciences,2018,19(10):E2883. [26] 赵嘉妮 ,陈宏 ,翁凌 ,等 .食源性 DPP-Ⅳ抑制肽降血糖作用机 制研究进展 [J].食品工业科技 ,2021,42(23):447-454. ZHAO J N,CHEN H,WENG L,et al. Research progress on the mechanism of food-derived DPP-Ⅳ inhibitory peptides for hypo-glycemic effect [ J]. Science and Technology of Food Industry,2021,42(23):447-454. (in Chinese) [27] XIE Y Z,LIU C H,CHENG L X,et al. Single molecular insight into steric effect on C-terminal amino acids with various hydrogen bonding sites [ J]. Chinese Chemical Letters, 2022, 33 ( 10 ):4649-4654. [28] JIN R T,TENG X Y,SHANG J Q, et all Identification of novel DPP-Ⅳ inhibitory peptides from Atlantic salmon ( Salmo salar ) skin[J]. Food Research International,2020,133:109-161. [29] 李艳敏 ,郁书怀 ,仝艳军 ,等 .裙带菜 α-葡萄糖苷酶抑制活性 肽的制备 [J].食品工业科技 ,2020,41(20):127-134. LI Y M,YU S H,TONG Y J,et al. Preparation of α-glucosidase inhibitory peptidesderived fromsea mustard [ J]. Scienceand Technology of Food Industry,2020,41( 20):127-134. ( in Chi-nese) [30] 袁娟 ,卫娜 ,徐勇 ,等 .白豆提取物对 α-淀粉酶和 α-葡萄糖苷 酶活性研究 [J].食品与发酵科技 ,2014,50(3):12-15. YUAN J,WEI N,XU Y,et al Research on α-amylase and α-glu-cosidase inhibitory activities of white beans natural extracts[J]. Food andFermentation Technology, 2014, 50 ( 3): 12-15. ( in Chinese) Structural characterization and the synergistic effects with anthocyanin of oyster hypoglycemic peptides JIANG Meiling1, CHEN Zhongqin1,2∗, QIN Xiaoming1,3∗, SUN Xujia1, CAO Wenhong1,2,3, LIN Haisheng1,3, GAO Jialong1,3 (1. National Research and Development Branch Center for Shellfish Processing (Zhanjiang), Guangdong Provincial Key Laboratory of Aquatic Products Processing and Safety, Guangdong Provincial Engineering Technology Research Center of Seafood, Guangdong Province Engineering Laboratory for Ma-rine Biological Products, Key Laboratory of Advanced Processing of Aquatic Product of Guangdong Higher Education Institution, College of Food Science and Technology, Guangdong Ocean University, Zhanjiang 524088, China; 2, Shenzhen Institute of Guangdong Ocean University, Shenzhen 518108, China; 3. Collaborative Innovation Center of Seafood Deep Processing, Dalian Polytechnic University, Dalian 116034, China) Abstract : To investigate the hypoglycemic effects and digestive stability of oyster peptides, the oyster peptides were prepared from Hong Kong oyster (Crassostrea hongkongensi )dStructural characteristics, hypoglycemic activity and digestive stability of prepared oyster peptides were investigated in vitro by hypoglycemic model and simulated gastro-intestinal digestion model-The anthocyanin was also used to improve the digestive stability and hypoglycemic activi-ty of prepared oyster peptidesoThe results showed that the prepared oyster peptides were rich in hydrophobic amino acids, and the relative molecular mass was mainlylessthan 1 000 The spectrum analysis showedthat prepared oyster peptides contained 15 peptide sequences mainly including LYF, TLFLK, IRAGYD, TLHHRVH, ARNEAN-VNIY, and CVIGR, which had typical structure characteristics of hypoglycemic peptides-The half inhibition con-centration (IC50)of prepared oyster peptides against α-amylase, α-glucosidase and dipeptidyl peptidase Ⅳ (DPP-Ⅳ)was (3.66±0.47), (8. 62±0) 66)and (2. 60±0. 46)mg/mL,respectively, exhibitinggood hypoglycemic activity However, the digestive stability of prepared oyster peptides was weak- After simulated gastrointestinal di-gestion in vitro ,the inhibition rate against the above three enzymessignificantly decreased by 6. 23%-32.48%(P <0 05) Therefore, the synergistic effects ofanthocyanin with preparedoyster peptides wereinvestigated Through the synergy matrix design, the highest single agent (HSA)model was selected to calculate the synergistic effects of anthocyanin with preparedoyster peptidesn The results showed that theoyster peptideand anthocyanin complex had significant synergistic inhibitory effects against α-amylase (HSA synergistic score 3.632), α-glucosi-dase (HSA synergistic score 3.503)and DPP-Ⅳ (HSA synergistic score 3.156)-Moreover, the results of simula-ted gastrointestinal digestion experiment in vitro showed that the anthocyanin could improve the digestive stability of prepared oyster peptidesf The results of this study indicate that oyster peptides have the characteristic structure of hypoglycemic peptides, and oyster peptides and anthocyanins have synergistic effect in enhancing hypoglycemic ac-tivity and digestive stability. Key words : oyster peptide; structural characterization; hypoglycemic activity; anthocyanin; synergistic effect
确定







还剩8页未读,是否继续阅读?

产品配置单







中国格哈特为您提供《牡蛎(生蚝)蛋白质含量的检测》,该方案主要用于生食水产品中营养成分检测,参考标准《GB 5009.5 食品安全国家标准 食品中蛋白质的测定》,《牡蛎(生蚝)蛋白质含量的检测》用到的仪器有格哈特全自动凯氏定氮仪VAPODEST 450、格哈特快速干燥仪STL56、格哈特红外加热消解快速消化系统TTs125、格哈特强力高重现振荡器LS500/RO500、格哈特维克松废气废物处理涤气系统VS、德国移液器MM、凯氏定氮催化剂5.0g K2SO4+0.5g CuSO4 x 5H2O
相关方案
更多
该厂商其他方案
更多